Opprinnelsen til kroppsdannelse
18.10.2013 06:52
Davidsons funn presenterer en dyptgående utfordring for tilstrekkeligheten til den neo-Darwinistiske mekanismen. Det å danne en ny type kropp krever ikke bare gener og proteiner, men nye dGRNs. Men en kan ikke lage nye dGRNs uten å endre eksisterende. Men det er nettopp dette som ikke er mulig, utenom
Fra 'Darwin's doubt' (Kap. 14)
 I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre kroppsplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..
I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre kroppsplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..
 30 år senere vet fortsatt ikke utviklings -og evolusjons-biologien svaret på det spørsmålet. På samme tid har mutasjonsgenetiske forsøk reist brysomme spørsmål angående rollen til mutasjoner i opphavet til utforming av dyrekropper. Det gjelder for forsøk på en rekke små dyrearter. Den neo-darwinistiske mekanismen har feilet i å forklare opphavet til nye gener og proteiner, nødvendig for at nye organer/kroppsdeler kan oppstå. Men selv om de kunne det, gjenstår et mye mer fundamentalt problem: For å bygge et nytt dyr og etablere kroppsplanen for det, trengs at proteiner arrangeres i høyere ordens strukturer. Med en gang nye proteiner oppstår, må 'noe' arrangere dem til å ta sin del i distinkte celle-typer. Disse må igjen organiseres, for å danne distinkte cellevev, organer og kropps-planer. Denne organiserings-prosessen foregår under utviklingen av embryos.
30 år senere vet fortsatt ikke utviklings -og evolusjons-biologien svaret på det spørsmålet. På samme tid har mutasjonsgenetiske forsøk reist brysomme spørsmål angående rollen til mutasjoner i opphavet til utforming av dyrekropper. Det gjelder for forsøk på en rekke små dyrearter. Den neo-darwinistiske mekanismen har feilet i å forklare opphavet til nye gener og proteiner, nødvendig for at nye organer/kroppsdeler kan oppstå. Men selv om de kunne det, gjenstår et mye mer fundamentalt problem: For å bygge et nytt dyr og etablere kroppsplanen for det, trengs at proteiner arrangeres i høyere ordens strukturer. Med en gang nye proteiner oppstår, må 'noe' arrangere dem til å ta sin del i distinkte celle-typer. Disse må igjen organiseres, for å danne distinkte cellevev, organer og kropps-planer. Denne organiserings-prosessen foregår under utviklingen av embryos.
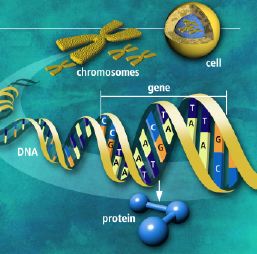 Utviklingsbiologien kaller prosessen der embryoet utvikler seg til modne organismer (gjelder flercellede organismer) for ontogeni. Feltet har stadig økt vår forståelse for hvordan kroppsplaner oppstår. Mye av kunnskapen har kommet gjennom såkalte modell-systemer, organismer som biologer lett kan mutere i laben (bananfluen Drosophila, samt en rundorm etc.) Selv om det er forvirrende ulikheter, er det ett felles trekk med all ontogeni: Den starter med ei celle og slutter med mange ulike celler. I de fleste arter starter det med ett befruktet egg. Med en gang egget deler seg i dets datter-celler, blir egget et embryo. Da begynner det å stile mot et vel-definert mål, nemlig en voksen form som kan reprodusere. For å nå det målet må embryoet produsere mange spesialiserte celle-typer på korrekt sted og tid.
Utviklingsbiologien kaller prosessen der embryoet utvikler seg til modne organismer (gjelder flercellede organismer) for ontogeni. Feltet har stadig økt vår forståelse for hvordan kroppsplaner oppstår. Mye av kunnskapen har kommet gjennom såkalte modell-systemer, organismer som biologer lett kan mutere i laben (bananfluen Drosophila, samt en rundorm etc.) Selv om det er forvirrende ulikheter, er det ett felles trekk med all ontogeni: Den starter med ei celle og slutter med mange ulike celler. I de fleste arter starter det med ett befruktet egg. Med en gang egget deler seg i dets datter-celler, blir egget et embryo. Da begynner det å stile mot et vel-definert mål, nemlig en voksen form som kan reprodusere. For å nå det målet må embryoet produsere mange spesialiserte celle-typer på korrekt sted og tid.
 Under ontogenien dobles antall celler hele tiden, ofte i en rasende fart. Antall celle-delinger og totalt antall celler reflekterer tallet på ulike celle-typer den modne organismen trenger. Dette innebærer igjen å produsere ulike proteiner for ulike celle-typer. Hvert protein må differensieres, fordi de gjerne utfører helt ulike oppgaver. Under ontogenien må da de rette genene bli slått av/på for å sikre at rett protein blir produsert til rett tid i de rette celle-typer. Spesifikke proteiner spiller aktive roller i regulering hvordan gener framtrer for å bygge andre proteiner. Proteinene som spiller denne rollen er kjent som transkripsjons-regulatorer(TRs) eller transkripsjons-faktorer (TFs). Disse knytter seg direkte til spesifikke steder i DNA og enten hindrer eller aktiver transkripsjon av spesifikke gener inn i RNA. De inneholder instruksjoner om hvilke gener de skal slå av eller på. Den tredimensjonale geometrien utøver karakteristiske trekk ved DNA-binding: inkl. et område av 61 aminosyrer som pakker seg omkring den doble DNA-spiralen.
Under ontogenien dobles antall celler hele tiden, ofte i en rasende fart. Antall celle-delinger og totalt antall celler reflekterer tallet på ulike celle-typer den modne organismen trenger. Dette innebærer igjen å produsere ulike proteiner for ulike celle-typer. Hvert protein må differensieres, fordi de gjerne utfører helt ulike oppgaver. Under ontogenien må da de rette genene bli slått av/på for å sikre at rett protein blir produsert til rett tid i de rette celle-typer. Spesifikke proteiner spiller aktive roller i regulering hvordan gener framtrer for å bygge andre proteiner. Proteinene som spiller denne rollen er kjent som transkripsjons-regulatorer(TRs) eller transkripsjons-faktorer (TFs). Disse knytter seg direkte til spesifikke steder i DNA og enten hindrer eller aktiver transkripsjon av spesifikke gener inn i RNA. De inneholder instruksjoner om hvilke gener de skal slå av eller på. Den tredimensjonale geometrien utøver karakteristiske trekk ved DNA-binding: inkl. et område av 61 aminosyrer som pakker seg omkring den doble DNA-spiralen.
 Transkripsjons regulatorer og faktorer (TRs og TFs) blir selv kontrollert av komplekse kretser og signaler, overført av andre gener og proteiner. Alt har en overveldene kompleksitet og presisjon. Mange av de regulerende gener som spiller nøkkel-roller i å omforme embryoer til modne organismer, er avdekket av forskningen til Wieshaus og Nüsslein-Vollhard. Denne forskningen avslørte også en fundamental vanskelighet, som gikk rett til 'hjertet' på det neo-Darwinistiske synet på livet.
Transkripsjons regulatorer og faktorer (TRs og TFs) blir selv kontrollert av komplekse kretser og signaler, overført av andre gener og proteiner. Alt har en overveldene kompleksitet og presisjon. Mange av de regulerende gener som spiller nøkkel-roller i å omforme embryoer til modne organismer, er avdekket av forskningen til Wieshaus og Nüsslein-Vollhard. Denne forskningen avslørte også en fundamental vanskelighet, som gikk rett til 'hjertet' på det neo-Darwinistiske synet på livet.
Mutasjoner tidlig i embryo-stadie
Normal utvikling for alle dyr kan representeres som et ekspanderende beslutnings-nettverk der de tidligste hendelser har større betydning enn senere. Regulerende gener og deres DNA-bindende protein kontrollerer dette utfoldende nettverket. Hvis da ett regulerende protein blir endret eller ødelagt av mutasjoner, så sprer effekten seg nedover i hele nettverket. Vanskeligheten med å forsøke endre en organisme til en annen, ligger i vanskeligheten ved å resette et antall av de mange kontrollerende svitsjene, slik at det tillater individets kroppsutvikling-uten å ødelegge det.
 Det bare mutasjoner framkalt tidlig i embryo-stadiet som har noen sjanse til å avstedkomme storskala, makroevolusjonær forandring. Sentvirkende mutasjoner kan dermed ikke framkalle store endringer. Dette bekreftes av flere evolusjonær-biologer (G. Miklos, K.Thompson ) Likevel i eksperimenter fra tidlige på 1900-tallet (T.H.Morgan..), så har erfaringen ensidig vært ødeleggende for organismer utsatt for mutasjon, inntil i dag.. Slike mutasjoner er enten dødelige eller resulterer i organismer som ikke kan leve i vill tilstand (neo-darwinist pioner R.A.Fisher) En endring tidlig i embryo-stadiet, ville kreve en rekke andre koordinerte forandringer i atskilte, men funksjonelt interrelaterte utviklingsprosesser. Det er den tette integreringen av fuksjoner som gjør at tidlige mutasjoner fører til deformering eller død i embryo-stadiet.
Det bare mutasjoner framkalt tidlig i embryo-stadiet som har noen sjanse til å avstedkomme storskala, makroevolusjonær forandring. Sentvirkende mutasjoner kan dermed ikke framkalle store endringer. Dette bekreftes av flere evolusjonær-biologer (G. Miklos, K.Thompson ) Likevel i eksperimenter fra tidlige på 1900-tallet (T.H.Morgan..), så har erfaringen ensidig vært ødeleggende for organismer utsatt for mutasjon, inntil i dag.. Slike mutasjoner er enten dødelige eller resulterer i organismer som ikke kan leve i vill tilstand (neo-darwinist pioner R.A.Fisher) En endring tidlig i embryo-stadiet, ville kreve en rekke andre koordinerte forandringer i atskilte, men funksjonelt interrelaterte utviklingsprosesser. Det er den tette integreringen av fuksjoner som gjør at tidlige mutasjoner fører til deformering eller død i embryo-stadiet.
dGRNs (development Gen Regulatory Networks)
 En annen linje i utviklings biologi har åpenbart en relatert utfordring til den kreative kraften i den neo-Darwinske mekanismen. Utviklingsbiologer har oppdaget at proteiner og RNA-er, som trengs for utvikling av kropps-utformingen til dyr, overfører signaler som influerer måten celler utvikler og differensieres. Disse signal-molekylene influerer hverandre til å danne nettverk av koordinert interaksjon (sml integrerte kretser på et kretskort). For eks. avhenger utskilling av signal-molekyl av når et signal mottas fra et annet molekyl. Og så påvirker det overføringen av enda flere, som alle er koordinerte og integrerte for å utførte spesifikke tids-kritiske funksjoner.
En annen linje i utviklings biologi har åpenbart en relatert utfordring til den kreative kraften i den neo-Darwinske mekanismen. Utviklingsbiologer har oppdaget at proteiner og RNA-er, som trengs for utvikling av kropps-utformingen til dyr, overfører signaler som influerer måten celler utvikler og differensieres. Disse signal-molekylene influerer hverandre til å danne nettverk av koordinert interaksjon (sml integrerte kretser på et kretskort). For eks. avhenger utskilling av signal-molekyl av når et signal mottas fra et annet molekyl. Og så påvirker det overføringen av enda flere, som alle er koordinerte og integrerte for å utførte spesifikke tids-kritiske funksjoner.
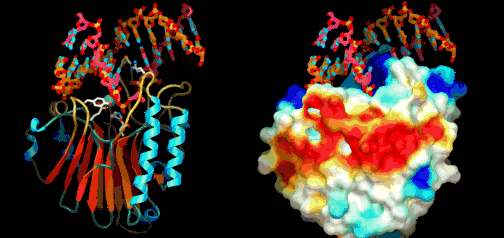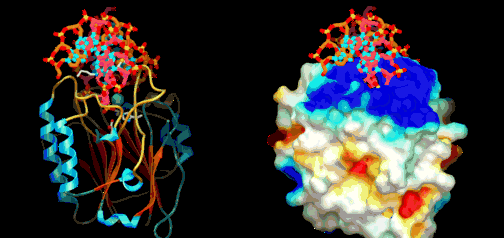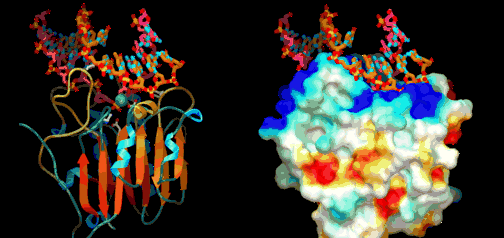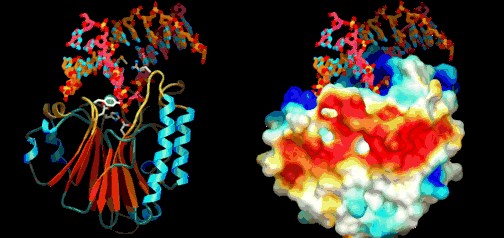 Eric Davidson er den biologen som mest dyptgående har undersøkt regulerings-logikken i dyrs utvikling. Davidson observerte at cellene til et individuelt dyr, samme hvor variert i form eller funksjon, generelt besto av identiske genomer. Gjennom livssyklusen til en organisme, så uttrykte genomet til disse spesialiserte cellene bare en liten del av sitt DNA til en gitt tid, og produserte dermed ulikt RNA som resultat. Dette faktum antyder sterkt at det finnes ett genetisk kontrollsystem for hele organismen, som skrur brytere på og av etter hvor en er i embryo-utviklingen. Likeså at et slikt system styrer utviklingen fra egg til moden organisme, i det ulike celle typer dannes. Britten og Davidson la fram en teori ang. dette i 1969. Selv om lite da var kjent, deduserte de at et slikt system måtte være virksomt. Det var for det første fordi hundrevis av spesialiserte celler oppsto gjennom utviklingen av dyr fra embryo til fullt utviklet. For det andre at hver celle inneholdt samme genom, så sluttet de at et slags kontroll-system måtte bestemme hvilke gener som uttrykkes i ulike celler til ulike tider, for å sikre differensiering av ulike celletyper fra hverandre. Derfor måtte det være en regulerende logikk som overstyrte og koordinerte hvordan genomet uttrykte seg til enhver tid.
Eric Davidson er den biologen som mest dyptgående har undersøkt regulerings-logikken i dyrs utvikling. Davidson observerte at cellene til et individuelt dyr, samme hvor variert i form eller funksjon, generelt besto av identiske genomer. Gjennom livssyklusen til en organisme, så uttrykte genomet til disse spesialiserte cellene bare en liten del av sitt DNA til en gitt tid, og produserte dermed ulikt RNA som resultat. Dette faktum antyder sterkt at det finnes ett genetisk kontrollsystem for hele organismen, som skrur brytere på og av etter hvor en er i embryo-utviklingen. Likeså at et slikt system styrer utviklingen fra egg til moden organisme, i det ulike celle typer dannes. Britten og Davidson la fram en teori ang. dette i 1969. Selv om lite da var kjent, deduserte de at et slikt system måtte være virksomt. Det var for det første fordi hundrevis av spesialiserte celler oppsto gjennom utviklingen av dyr fra embryo til fullt utviklet. For det andre at hver celle inneholdt samme genom, så sluttet de at et slags kontroll-system måtte bestemme hvilke gener som uttrykkes i ulike celler til ulike tider, for å sikre differensiering av ulike celletyper fra hverandre. Derfor måtte det være en regulerende logikk som overstyrte og koordinerte hvordan genomet uttrykte seg til enhver tid.
 Gjennom de siste to tiår har forskning i genomet avslørt at ikke-kodende regioner av genomet kontrollerer og regulerer timingen til hvordan protein-kodende regioner av genomet uttrykker seg. Sammen fungerer ikke-protein kodende og protein-kodende regioner av genomet som et nettverk i følge Davidson. Davidson valgte fra 1971 et sjøpiggsvin (Strongylocentrotus purpuratus) som modell for sitt eksperiment. Sammen med kolleger utviklet Davidson et pioner-prosjekt og eksperimentelle protokoller som krevdes for å dissekere og kartlegge sjøpiggsvinets genetiske regulerende nettverk.
Gjennom de siste to tiår har forskning i genomet avslørt at ikke-kodende regioner av genomet kontrollerer og regulerer timingen til hvordan protein-kodende regioner av genomet uttrykker seg. Sammen fungerer ikke-protein kodende og protein-kodende regioner av genomet som et nettverk i følge Davidson. Davidson valgte fra 1971 et sjøpiggsvin (Strongylocentrotus purpuratus) som modell for sitt eksperiment. Sammen med kolleger utviklet Davidson et pioner-prosjekt og eksperimentelle protokoller som krevdes for å dissekere og kartlegge sjøpiggsvinets genetiske regulerende nettverk.
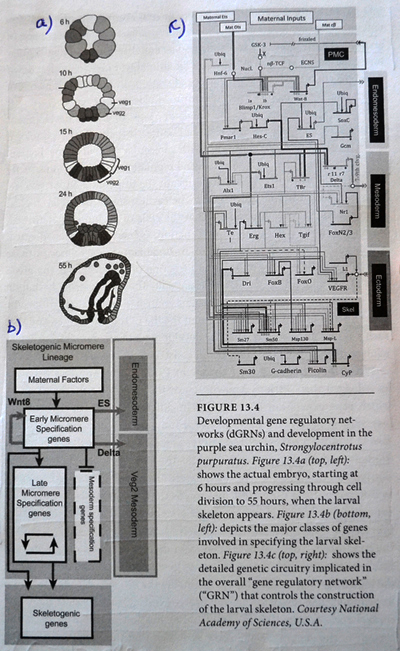 Den fantastiske kompleksiteten de fant framstilles på figure 13.4 her ved siden. Del a viser utvikling av embryoet i en alder fra 6 til 55 timer. Øverst er det 4 runder med celledeling og 16 celler (2 opphøyd i 4). I de neste fire stadiene ser vi en tiltagende spesialisering i kroppformen. Del b er et skjematisk diagram over hovedklassene av gener og cellevev i løpet av embryo-utviklingen. Sammenhengen markeres med kontrollpiler. Fig 13.c viser 'det genetiske kretskortet' (i følge Davidson) som skrur på de spesifikke genene som produserer det nødvendige strukturelle proteinet for å danne sjøpiggsvinet.
Den fantastiske kompleksiteten de fant framstilles på figure 13.4 her ved siden. Del a viser utvikling av embryoet i en alder fra 6 til 55 timer. Øverst er det 4 runder med celledeling og 16 celler (2 opphøyd i 4). I de neste fire stadiene ser vi en tiltagende spesialisering i kroppformen. Del b er et skjematisk diagram over hovedklassene av gener og cellevev i løpet av embryo-utviklingen. Sammenhengen markeres med kontrollpiler. Fig 13.c viser 'det genetiske kretskortet' (i følge Davidson) som skrur på de spesifikke genene som produserer det nødvendige strukturelle proteinet for å danne sjøpiggsvinet.
For å uttrykke genene som lager proteiner for å danne skjelettet, så må først gener som aktiveres timer tidligere først spille sin rolle. Denne prosessen skjer ikke på slump, men via høyst regulerte og presise kontrollsystemer som det gjør i alle alle dyr. Tiden til rådighet for mutasjoner er heller knapp, for enklere organismer: noen få døgn. De enkleste organismer, (f.eks. en voksen sjøorm), har kun litt over 1000 celler. Likevel har den dGRNs av bemerkelsesverdig presisjon og kompleksitet. Utviklingen av kompleksiteten i embryoet kan måles i informasjonstermer, i følge Davidson.
 Developmental gen regulatory networks (Dgrns) motstår mutasjonsendringer fordi de er hierarkisk organisert. Det innebærer at noen Dgrns kontrollerer andre Dgrns. I senteret av dette regulerende hierarkiet, er regulerende nettverk som spesifiserer akse og helhetlig kroppsplan under utviklingen. Selv små endringer i disse Dgrns vil medføre katastrofale virkninger på organismen. Likevel er det nettopp det som trengs om akse og global form på dyret skal variere: Nettopp de kretsløpene som ikke kan endres uten ødeleggelse som følge, må gjøre det om kroppsformen skal endres.
Developmental gen regulatory networks (Dgrns) motstår mutasjonsendringer fordi de er hierarkisk organisert. Det innebærer at noen Dgrns kontrollerer andre Dgrns. I senteret av dette regulerende hierarkiet, er regulerende nettverk som spesifiserer akse og helhetlig kroppsplan under utviklingen. Selv små endringer i disse Dgrns vil medføre katastrofale virkninger på organismen. Likevel er det nettopp det som trengs om akse og global form på dyret skal variere: Nettopp de kretsløpene som ikke kan endres uten ødeleggelse som følge, må gjøre det om kroppsformen skal endres. multippelt koordinerte mutasjoner. I et hvert tilfelle har Davidsons arbeid vist oss at slike endringer alltid har ført til katastrofale konsekvenser. Han gjør dette helt klart: 'I motsetning til klassisk evolusjonsteori, så kan ikke prosesser som drives fram av små endringer bli tatt som modell for utviklingen av kropps-form for dyrearter'.. Dette kan ikke være overraskende, i følge Davidson, da neo-Darwinistisk syntese, stammer fra en premolekylær biologi fokusert på populasjons genetikk og ..naturlig historie. Ingen av disse har noen inngang for dGRNs som styrer embryo-utvikling av kroppsformer..
multippelt koordinerte mutasjoner. I et hvert tilfelle har Davidsons arbeid vist oss at slike endringer alltid har ført til katastrofale konsekvenser. Han gjør dette helt klart: 'I motsetning til klassisk evolusjonsteori, så kan ikke prosesser som drives fram av små endringer bli tatt som modell for utviklingen av kropps-form for dyrearter'.. Dette kan ikke være overraskende, i følge Davidson, da neo-Darwinistisk syntese, stammer fra en premolekylær biologi fokusert på populasjons genetikk og ..naturlig historie. Ingen av disse har noen inngang for dGRNs som styrer embryo-utvikling av kroppsformer..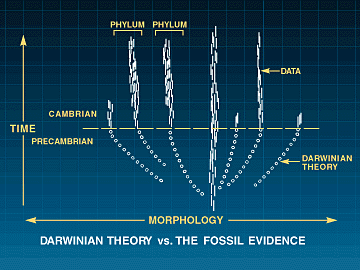 Paleontologer forstår den cambriske eksplosjonen som en plutselig geologisk tilsynekomst av nye livsformer. Å bygge disse kroppsuttrykkene krever nye utviklings program, både nye tidlig-handlende regulerende gener OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet om å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre. Ennå gjenstår mer formidable problemer for neo-Darwinismen (Kap.14-epigenetikk).
Paleontologer forstår den cambriske eksplosjonen som en plutselig geologisk tilsynekomst av nye livsformer. Å bygge disse kroppsuttrykkene krever nye utviklings program, både nye tidlig-handlende regulerende gener OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet om å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre. Ennå gjenstår mer formidable problemer for neo-Darwinismen (Kap.14-epigenetikk).