Homologi i virveldyrs lemmer
23.11.2013 10:46
Hvis virveldyrs utvikling av lemmer skulle avspeile en felles opprinnelse, måtte en forvente at en kan iaktta det samme nedarvede bruskmønsteret tidlig i virveldyrs utvikling av lemmer. Men slik er det ikke: Bruskmønsteret svarer fra første stund til den form det voksne individ har, ikke bare hos salamandere, men også hos padder, høns og mus. Oppfattelsen av at lemmer skulle komme fra et felles nedarvet mønster i embryoet, er i følge de britiske zoologer Richard Hinchlife og P.J.Griffiths "oppstått fordi forskerne har lagt sine forut-innfattede meninger" ned over forskningsresultatene.
(Fra 'Evolutionens ikoner'; Jonathan Wells; Forlaget Origo)
Siden den greske filosofen Aristoteles i antikken har det vært lagt merke til at ulike organismer kan ha visse likheter. Den ene form for likhet er funksjonell: både sommerfugler og flaggermus har vinger til å fly med. De to dyrene er konstruert rimelig forskjellig. En annen form for likhet er strukturell. Knokkelstrukturen i en flaggermusvinge ligner den som finnes i en sels luffe, selv om den første brukes til å fly med og luffen til å svømme med. Vitenskapelig sett kommer vi da til morfologi, som er læren om former (gresk lógos = «lære», morphé = «form»). I biologien refererer det til kroppsbygningen hos en organisme og faget som studerer dette.
 I 1840 årene kalte den britiske anatom Richard Owen den første form likheter for 'analogi' og den annen form for 'homologi'. Den gang ble skjelningen primært anvendt som hjelp til biologisk klassifisering. Analogi kan tyde på uavhengige tilpasninger til ytre forhold, mens homologi kan tyde på dypere strukturelt slektskap. Sistnevnte ble regnet for å være mest pålitelig når det gjaldt gruppering av organismer i familier, ordener, klasser og rekker.
I 1840 årene kalte den britiske anatom Richard Owen den første form likheter for 'analogi' og den annen form for 'homologi'. Den gang ble skjelningen primært anvendt som hjelp til biologisk klassifisering. Analogi kan tyde på uavhengige tilpasninger til ytre forhold, mens homologi kan tyde på dypere strukturelt slektskap. Sistnevnte ble regnet for å være mest pålitelig når det gjaldt gruppering av organismer i familier, ordener, klasser og rekker.
Som mange andre pre-darwinistiske biologer mente Owen at homologe trekk nedstammer fra en felles 'arketype'. Det kan imidlertid forstås på ulike vis: -som en legemsløs, platonisk idé, en plan i Skaperens hode, en fra naturen iboende aristotelisk struktur eller prototypisk organisme osv. Både Owen og Darwin så arketypen som en prototypisk organisme. Owen mente at organismene var konstruert over en felles plan, en prototype. Mens Darwin senere kom til at de nedstammet fra en felles forfader.
I 'Artenes opprinnelse' argumenterte Darwin for at den beste forklaring på homologi er nedarving med tilpasning. 'Hvis vi går ut fra en tidlig stamfar -arketypen om en vil -til alle pattedyr, fugler og krypdyr, fikk sine lemmer konstruert etter den eksisterende arketypen så ville den samme skjelettstruktur i et menneskes hånd, en flaggermus sin vinge, en sels luffe og en hests ben plutselig kunne forklare seg selv etter teorien om nedarving med langsomme modifikasjoner.' Darwin så homologien som et viktig vitnesbyrd om evolusjonen og regnet det blant 'de kjennsgjerninger som så tydelig proklamerer at utallige arter, slekter og familier som befolker denne verden alle har de samme foreldre, enhver innenfor sin egen klasse eller gruppe.'
Nåtidens darwinister anvender fortsatt homologi som argument for sin teori. Nest etter Darwins evolusjonstre, er homologi i virveldyrs lemmer, sannsynligvis 'det ikon' en oftest støtter på i lærebøker i biologi. Imidlertid skjuler det seg to alvorlige problemstillinger i dette: i) Homologi er gått over til å bety 'likheter som skyldes felles avstamning' (Andre likheter kalles for 'analoge'.) Hvordan kan det 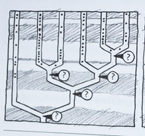 da benyttes til å argumentere for felles nedstamning. Det blir som å si at 'felles nedstamning skyldes felles nedstamning.' ii) Selv om det finnes enkelte felles gener med lik funksjonell virkning, har biologien mer blitt klar over at hver art har ulike epigenom, med kontekstavhengig informasjon for hver celletype. Dermed forblir mekanismen som skulle fremkalle homologi fortsatt ikke avklart!
da benyttes til å argumentere for felles nedstamning. Det blir som å si at 'felles nedstamning skyldes felles nedstamning.' ii) Selv om det finnes enkelte felles gener med lik funksjonell virkning, har biologien mer blitt klar over at hver art har ulike epigenom, med kontekstavhengig informasjon for hver celletype. Dermed forblir mekanismen som skulle fremkalle homologi fortsatt ikke avklart!
Som nevnt er begrepet homologi blitt redefinert: Før Darwin var definisjonen på homologi likheter i strukturer og plassering. For biologer før Darwin var forklaringen en avledning av et opprinnelig mønster eller en arketype. Darwin omdefinerte avledning til biologisk evolusjon og en arketype til en felles forfader. Ernst Mayr en av neo-darwinismens hovedarkitekter, sier det slik: "Etter 1859 har det kun vært én definisjon på begrepet homologi som gir biologisk mening..To organismers attributter er homologe når de begge er avledet fra ekvivalente kjennetegn hos en felles forfader.'
For Charles Darwin var utviklingslæren en teori, og homologi var et tegn på teoriens sannhet. For Darwins etterfølgere er utviklingslæren en velfundert sannhet, og homologien anses å være resultatet av denne. Problemet er at nå kan ikke homologi benyttes som argument for utviklingslæren, uten å gå i en sirkelslutning.
Homologi og sirkelargumentasjon
 La oss betrakte eksempelet med forlemmers knokkelstrukturer (figur ovenfor) som Darwin mente var et vitnesbyrd om felles avstamning. En neodarwinist som vil avgjøre om virveldyrs forlemmer er homologe, må først avgjøre om de stammer fra en felles forfader eller ikke. Det må med andre ord være belegg for et felles opphav, før en kan kalle lemmer for homologe. Men hvis en så vender argumentasjonen rundt og argumenterer for at homologe lemmer peker på felles avstamning, blir det en grov sirkelslutning: Et felles opphav viser homologi, som i sin tur viser felles opphav. (Fig.4.2)
La oss betrakte eksempelet med forlemmers knokkelstrukturer (figur ovenfor) som Darwin mente var et vitnesbyrd om felles avstamning. En neodarwinist som vil avgjøre om virveldyrs forlemmer er homologe, må først avgjøre om de stammer fra en felles forfader eller ikke. Det må med andre ord være belegg for et felles opphav, før en kan kalle lemmer for homologe. Men hvis en så vender argumentasjonen rundt og argumenterer for at homologe lemmer peker på felles avstamning, blir det en grov sirkelslutning: Et felles opphav viser homologi, som i sin tur viser felles opphav. (Fig.4.2)
Polemikken har pågått siden Darwin. Neo-darwinister forsvarer forestillingen om homologi som tegn på felles avstamning. Kritikere av dette synet hevder at en blander definisjon og konklusjon sammen. Filosofen Ronald Brady skrev i 1985: " Ved å trekke vår forklaring inn i definisjonen av de forhold som skal forklares, fremlegger vi ikke en vitenskapelig hypotese, men vi bringer en overbevisning til torvs. Vi er så overbevist om at vår forklaring er sann, at vi ikke lenger synes det er nødvendig å holde den atskilt fra det forhold vi prøver å forklare. Den slags dogmatiske forehavender må og skal vi ha utryddet i den vitenskapelige begrepsverden.'
Kan sirkelen brytes?
Tilsynelatende er det tre måter å unngå sirkeslutningen en kommer inn i om en både definerer og forklarer homologi ut fra felles avstamning. Den ene er å bruke homologi som definisjon på felles opphav, men unngå å utlede felles avstamning ut fra den. M.a.o. anerkjenne at homologi ikke lenger underbygger utviklingslæren. Evolusjonsbiolog David Wake skrev i 1999: Derfor "er homologi den forutsatte og forventede konsekvens av evolusjon. Homologi er ikke et vitnesbyrd om evolusjon."
Den andre måten er å gjeninnføre den pre-darwinistiske definisjon på homologi, som en strukturlikhet og samtidig anerkjenne at dette gjenåpner spørsmålet hvorvidt 'nedarving med tilpasning' er den beste  forklaring på fenomenet. I USA kan det være vanskelig å finne fortalere for denne retningen, fordi det blant biologer er meget upopulært og risikabelt profesjonelt å sette spørsmålstegn ved Darwinismen.
forklaring på fenomenet. I USA kan det være vanskelig å finne fortalere for denne retningen, fordi det blant biologer er meget upopulært og risikabelt profesjonelt å sette spørsmålstegn ved Darwinismen.
Den tredje og for tiden mest populære måte å løse problemet på er å definere homologi som uttrykk for felles avstamning, og derpå finne forskningsresultater som peker i retning av nedarving med tilpasning, uavhengig av homologien. Slike forskningsresultat kan være strukturorientert, f.eks. i form av DNA-sekvenssamlinger eller fossilmaterialet. Eller det kan være prosessorientert: utviklingsretninger eller utviklingsgenetikk. De to første legger ut med en antagelse om felles avstamning og førsøker dernest å utlede det mest sannsynlige mønster for de ulike slektskapsforhold. De to sistnevnte forsøker finne de prosesser som kan være ansvarlige for likheter som skyldes felles avstamning.
 Hva viser DNA-sekvenser i retning av
Hva viser DNA-sekvenser i retning av
Molekylær fylogenese (omhandler slektskapsforholdene mellom arter eller grupper av arter) bygges opp ved sammenlikning av DNA-sekvenser (eller deres proteinprodukter) i ulike organismer. DNA-sekvenser kopieres direkte fra andre DNA-sekvenser via replikasjonsprosessen. Derfor formoder molekylær-fylogenetikerne at sekvenslikheter nok i høyere grad enn morfologiske likheter peker på et evt. slektskapsforhold
Uheldigvis er det like så mange vansker ved molekylære sekvensanalyser, som ved morfologiske sammenligninger. Molekylærbiologen David Hills skrev i 1994: "Ordet homologi brukes nå innen biologien til å beskrive alt fra ordinære likheter til felles avstamning".. Således har "molekylærbiologer kanskje gjort mer for å forvirre om begrepet homologi enn noen anne gruppe av forskere."
Homologi på molekylærnivå skaper minst like mange motsigende resultater som i andre fag-grener. De britiske biologer Colin Patterson, David Williams og Christopher Humphries skrev i 1993: "Overensstemmelse mellom de molekylære fylogeneser er like så vanskelig å bestemme som de  er i morfologien." Så når de molekylære fylogeneser ikke stemmer overens, er eneste måteen kan skjelne mellom dem, at en vet noe om deres felles avstamning fra andre kilder -og herved er vi direkte tilbake i de sirkelslutninger som molekylære sammenlikninger skulle hjelpe oss komme ut av." (Fylogenese: utforsker slektskapsforholdene mellom arter eller grupper av arter. Ordet kommer av de greske røttene φύλον (fylon), «stamme» og γένεσις (genesis), «dannelse».)
er i morfologien." Så når de molekylære fylogeneser ikke stemmer overens, er eneste måteen kan skjelne mellom dem, at en vet noe om deres felles avstamning fra andre kilder -og herved er vi direkte tilbake i de sirkelslutninger som molekylære sammenlikninger skulle hjelpe oss komme ut av." (Fylogenese: utforsker slektskapsforholdene mellom arter eller grupper av arter. Ordet kommer av de greske røttene φύλον (fylon), «stamme» og γένεσις (genesis), «dannelse».)
Den fossile lagserie
Noen biologer har hevdet at en måte å fastsette evolusjonære avstamningsforhold på, er å spore likhetene i to eller flere organismer via en ubrutt kjede av fossile organismer, tilbake til sitt fossile opphav. Uheldigvis er det å sammenligne fossiler ikke mindre vanskelig enn å sammenligne levende prøveeksemplarer. Sokal og Sneath skrev i 1963: "Selv om det er fossilmateriale tilgjengelig, må dette materiale først fortolkes" ved sammenlikning av ensartede trekk. Ethvert forsøk på å dra slutninger om  fossilenes evolusjonære slektskap som bygger på homologi som begrunnelse for felles avstamning " fanger oss hurtig inn i et nett av sirkelslutninger som det er umulig å unnslippe."
fossilenes evolusjonære slektskap som bygger på homologi som begrunnelse for felles avstamning " fanger oss hurtig inn i et nett av sirkelslutninger som det er umulig å unnslippe."
Faktisk er det vanskeligere å utlede evolusjonært slektskap ut fra den fossile lagserie enn fra levende organismer, fordi fossilhistorien er fragmentarisk, og fossilene ikke har bevart alle relevante trekk. Biologen Bruce Yong skrev i 1993: "Hvis fossiler overhodet er noe, så er de i hvert fall av mindre verdi når man skal fastslå homologe forhold, ettersom de normalt omfatter langt færre egenskaper enn levende organismer."
Men selv om den fossile lagserie hadde vært fullstendig, og den hadde bevart alle ønskede egenskaper, ville den likevel ikke godtgjøre at homologi skyldes et felles opphav. Det finnes andre forklaringer enn nedarving ved tilpasning på morfologiske likheter. De fleste biologer før Darwin ville ha forklart slike likheter med felles design eller skapelse. Fossilmaterialet er forenlig med begge forklaringer. En enkel oppstilling av former som ligner hverandre, gir ikke en forklaring på opphavet i seg selv. Det kreves en mekanisme som kan forklare slike likheter. Forklaringer som "nedstamning og tilpasning" er bare ord, om de ikke kan kobles mot faktiske biologiske prosesser.
 Vidnesbyrd fra utviklingsretninger
Vidnesbyrd fra utviklingsretninger
Teorien om at homologe strukturer er resultat av sammenlignbare utviklingsretninger, passer ikke med tilstedeværende forskningsmateriale. Det har biologer visst i mer enn et århundre. Den amerikanske embryolog Edmund Wilson skrev i 1894: "at deler som stemmer nøye overens på voskenstadiet .. ofte er veldig forskjellige fra hverandre på larve eller fosterstadiet, både med hensyn til dannelse og/eller plassering." Etter å ha studert det embryologiske forskningsmaterialet som har hopet seg opp etter det, gav den britiske biolog Gavin de Beer ham rett -mer enn 60 år senere: "Faktum er at en sammenheng mellom homologe strukturer ikke kan tolkes ned over likheter i fostercellers plassering eller i de deler av egget der de første strukturer er dannet, eller i de utviklingsmekanismer som har dannet dem."
I 1985 skrev utviklingsbiologen Pere Alberch at"homologe strukturer har deres opprinnelse i klart forskjellige utgangspunkter." Forsker i evolusjonær utviklingsbiologi Rudolf Raff skrev i 1999: "Homologe trekk i to beslektede organismer burde oppstå ved homologe utviklingsprosesser..[men det er faktisk slik at]trekk vi ser som homologe ut fra morfologiske eller fyliogenetiske kriterier, kan oppstå på forskjellig vis i utviklingen [av individet]."
 Manglende sammenheng mellom homologi og utviklingsretninger, er gjeldende ikke bare i store trekk, men også i helt spesielle tilfeller ved virveldyrs lemmer. Det klassiske eksempel er salamanderne. I de fleste virveldyrs lemmer foregår utviklingen av fingre og tær bakfra og framover (fra hale mot hode), men hos salamanderne er det motsatt. Noen biologer har hevdet at salamandernes evolusjonshistorie må ha vært annerledes enn alle andre virveldyrs. Også når det gjelder utviklingen av virveldyrs skjelett-struktur, -som først formes som brusk og senere blir knokkelvev, er det uregelmessigheter:
Manglende sammenheng mellom homologi og utviklingsretninger, er gjeldende ikke bare i store trekk, men også i helt spesielle tilfeller ved virveldyrs lemmer. Det klassiske eksempel er salamanderne. I de fleste virveldyrs lemmer foregår utviklingen av fingre og tær bakfra og framover (fra hale mot hode), men hos salamanderne er det motsatt. Noen biologer har hevdet at salamandernes evolusjonshistorie må ha vært annerledes enn alle andre virveldyrs. Også når det gjelder utviklingen av virveldyrs skjelett-struktur, -som først formes som brusk og senere blir knokkelvev, er det uregelmessigheter:
Hvis virveldyrs utvikling av lemmer skulle avspeile en felles opprinnelse, måtte en forvente at en kan iaktta det samme nedarvede bruskmønsteret tidlig i virveldyrs utvikling av lemmer. Men slik er det ikke: Bruskmønsteret svarer fra første stund til den form det voksne individ har, ikke bare hos salamandere, men også hos padder, høns og mus. Oppfattelsen av at lemmer skulle komme fra et felles nedarvet mønster i embryoet, er i følge de britiske zoologer Richard Hinchlife og P.J.Griffiths "oppstått fordi forskerne har lagt sine forut-innfattede meninger" ned over forskningsresultatene.
Homologe trekk i virveldyrs lemmer er altså ikke framkommet av utviklingsretninger som svarer til hverandre. Hva så med genene?
Vitnesbyrd fra utviklingsgenetikken
 I følge neodarwinismen er det DNA-sekvenser eller gener som inneholder informasjon. Gener overfører informasjon fra generasjon til generasjon og styrer, i følge teorien, fosterets utvikling. Derfor er den teoretiske forklaring på homologe trekk at de er programmert av ensartede gener, nedarvet fra en felles forfader. Hvis en kan påvise at homologe strukturer i to forskjellige organismer er framkalt av tilsvarende gener, så ville vi ha forskningsmessig belegg for "informasjonskontinuiteten".
I følge neodarwinismen er det DNA-sekvenser eller gener som inneholder informasjon. Gener overfører informasjon fra generasjon til generasjon og styrer, i følge teorien, fosterets utvikling. Derfor er den teoretiske forklaring på homologe trekk at de er programmert av ensartede gener, nedarvet fra en felles forfader. Hvis en kan påvise at homologe strukturer i to forskjellige organismer er framkalt av tilsvarende gener, så ville vi ha forskningsmessig belegg for "informasjonskontinuiteten".
Men slik forholder det seg ikke. Det har biologer kjent til i årtier. I 1971 skrev Gavin de Beer: "Ettersom homologi forutsetter et nedstamningsforhold fra en felles forfader, kunne en tro at genetikken ville stå med nøkkelen til homologiproblemet. Det er her vi støter på det verst tenkelige sjokk av alle..[fordi] legemstrekk som kontrolleres av ensartede gener ikke nødvendigvis er homologe..[og] homologe strukturer ikke nødvendigvis kontrolleres av ensartede gener." De Beer konkluderer at "nedarvingen av homologe strukturer fra en felles forfader .. kan ikke tilskrives identiske gener".
 Som eks. på dette anførte de Beer kun ét eksperiment som har med øyeutvikling hos bananfluen. Siden har en støtt på andre eksempler. Ett av dem er insekters leddannelse. Bananflue krever genet 'even-skipped' til utvikling av kroppsdeler, mens andre insekter utvikler ledd uten dette genet. Da alle insekt-ledd ansees være homologe, viser dette eksempel at homologe trekk ikke trenger være styrt av ensartede gener. Et annet eks. er 'sex-lethal', et gen som kreves til bestemmelse av kjønnet hos bananfluer, men ikke hos andre insekter..
Som eks. på dette anførte de Beer kun ét eksperiment som har med øyeutvikling hos bananfluen. Siden har en støtt på andre eksempler. Ett av dem er insekters leddannelse. Bananflue krever genet 'even-skipped' til utvikling av kroppsdeler, mens andre insekter utvikler ledd uten dette genet. Da alle insekt-ledd ansees være homologe, viser dette eksempel at homologe trekk ikke trenger være styrt av ensartede gener. Et annet eks. er 'sex-lethal', et gen som kreves til bestemmelse av kjønnet hos bananfluer, men ikke hos andre insekter..
Den omvendte situasjon, at ikke-homologe strukturer oppstår fra identiske gener, er både mer bemerkelsesverdig og mer utbredt. Genetikere har funnet at mange av de gener som kreves til bananfluers utvikling, svarer til de gener som finnes i mus, sjøpiggsvin og mark. Det er faktisk slik at gentransplanasjoner har vist at utviklingsgener fra mus kan erstattes av deres motstykker hos fluer. Men hvis disse genene ligner hverandre så mye, hvorfor utvikler så ikke et musefoster seg til å bli ei flue, eller vice versa?
 Mangel på overensstemmelse mellom gener og strukturer gjelder ikke bare for hele organismer, men også for lemmer. Et utviklingsgen som finnes i alle organismer, kalles 'Distal-less'. Det har fått dette navnet fordi en mutasjon i dette gen blokkerer bananfluers utvikling av lemmer (distal vedrører strukturer som ikke er en del av selve kroppen). Et gen med en ganske tilsvarende DNA-sekvens er funnet husmus. Gener som ligner 'Distal-less' er også funnet i sjøpiggsvin, børsteorm og ledd-orm lignende dyr (Onychophora).
Mangel på overensstemmelse mellom gener og strukturer gjelder ikke bare for hele organismer, men også for lemmer. Et utviklingsgen som finnes i alle organismer, kalles 'Distal-less'. Det har fått dette navnet fordi en mutasjon i dette gen blokkerer bananfluers utvikling av lemmer (distal vedrører strukturer som ikke er en del av selve kroppen). Et gen med en ganske tilsvarende DNA-sekvens er funnet husmus. Gener som ligner 'Distal-less' er også funnet i sjøpiggsvin, børsteorm og ledd-orm lignende dyr (Onychophora).
Hos alle disse dyreartene er 'Distal-less' forbundet med utviklingen av deres vedheng (ben), men benene er ikke homologe, verken på struktur- eller på utviklingsnivå. Dette synes forbløffende, fordi vedhengene har så ulik anatomi og utviklingshistorie. I 1999 konkluderer Gregory Wray: "Denne sammenhengen mellom ett regulerende gen og atskillige ikke-homologe strukturer ser snarere ut til å være regelen enn unntaket." Hele det nettverk av gener som er knyttet til lemmers utvikling, ligner hverandre hos både insekter og virveldyr. Clifford Tabin, Sean Carroll og Grace Panganiban skrev i 1999 at: "der ikke har vært noen sammenheng mellom noen av de strukturer hvorfra insekters og virveldyrs vedheng kan utledes"; altså er de ikke homologe ( i darwinistisk betydning). Allikevel tyder mye på at det er sammenheng mellom den genetiske informasjonen som står for deres utvikling.
Evolusjonsbiologer hevder at den slående likheten mellom utviklingsgener i en så bred vifte av dyrearter, tyder på et felles opphav. Det kan det også godt være, på annet vis enn de tenker.. For dem dukker problemene vi møtte i den molekylære fylogenesen opp igjen, mens hele problematikken med å forklare homologe strukturers opprinnelse forblir uløst.
Konklusjonen står klar: Enten homologi skyldes nedarving med tilpasning eller ikke, forblir den spesifikke mekanismen bak det hele ukjent. Gavin de Beer skrev i 1971: "Hvilken mekanisme kan resultere i framstillingen av homologe organer, tross at de ikke styres av de samme gener? Jeg har stilt dette spørsmålet i 1938 og jeg har ennå ikke fått svar på det." Dette spørsmålet står fortsatt ubesvart.
Virveldyrs lemmer som vitnesbyrd om evolusjon
På hvilken måte kan virveldyrs lemmer utgjøre et vitnesbyrd og Darwins utviklingslære? Hvis informasjonskontinuiteten ikke kommer fra gener eller utviklingsretninger, hvordan kan vi vite at det skyldes nedarving med tilpasning? Er det mulig å utlede felles avstamning fra homologi på annen måte enn å definere det inn i forutsetningene? 'Hvis definisjonen på  homologi er felles avstamning, kan ikke homologi brukes som vitnesbyrd om felles avstamning'. Lærebøker i biologi prøver å underslå sirkelslutningen de gjør. Det er et problem for demokratiet. Samfunnet har bruk for folk som kan tenke selv, og ikke bare føyer seg etter hva eksperter og autoriteter forer dem med..
homologi er felles avstamning, kan ikke homologi brukes som vitnesbyrd om felles avstamning'. Lærebøker i biologi prøver å underslå sirkelslutningen de gjør. Det er et problem for demokratiet. Samfunnet har bruk for folk som kan tenke selv, og ikke bare føyer seg etter hva eksperter og autoriteter forer dem med..